| цкюбмюъ | днйкюдш | наясфдемхе | |
Направленность эволюции и регулирующие механизмы видообразования: кибернетический аспект
А.Р. Галеев
Кемеровский государственный университет
1. Введение
В настоящее время широкое распространение получил системный подход к анализу явлений природы и общества. В значительной мере это явилось необходимостью практики. Изучение таких глобальных проблем, как здоровье людей, экология, исследование мирового океана, космоса и т.п. потребовало применения системных подходов.
Пожалуй, одной из первых биологических наук, в которой интенсивно применялся системный подход, была эволюционная биология. Уже в середине 19 века в ней нашли применение данные и методы почти всех естественных наук. Следствием этого было два синтеза накопленных знаний: теория естественного отбора Ч. Дарвина и современная синтетическая теория эволюции. Недалек момент, когда накопление новых данных приведет к необходимости обновления и дополнения теории эволюции и нового синтеза.
Вместе с этим, удивительно, что на периферии эволюционного учения остается кибернетика. Кибернетика √ это наука об общих закономерностях процессов управления и связях в системах любой природы и сложности и об организации этих систем. Соответственно, предметом кибернетики являются законы и закономерности организации систем
. Поскольку биогеоценоз, популяция и организм представляют собой системы, то, очевидно, что кибернетика могла бы стать одним из связующих звеньев в многогранной эволюционной биологии.В результате развития кибернетики и пристального интереса к системному подходу была создана общая теория систем [1]. Сторонники данной теории рассматривают системы, как множество взаимосвязанных элементов. При этом системы обладают качественно иными свойствами по отношению к составляющим их элементам. Революционным введением в теорию систем было понятие ╚системообразующий фактор╩, сформулированное П.К. Анохиным [2]. Системообразующим фактором является полезный для деятельности системы приспособительный результат. В результате многочисленных исследований живых организмов П.К. Анохин разработал теория функциональных систем. Согласно теории функциональные системы организма √ это саморегулирующиеся динамические центрально-периферические построения, объединенные регуляцией, все составные компоненты которых взаимодействуют для обеспечения полезных адаптивных результатов, удовлетворяющие различные потребности, как отдельных систем организма, так и организма в целом. Оценка параметров достигнутых результатов в функциональной системе постоянно осуществляется с помощью обратной связи.
Очевидно, что методы и понятия, используемые в кибернетике и входящей в нее теории функциональных систем, могли бы найти приложение в толковании многих эволюционных явлений. Поскольку как сам организм, так и популяция (единица эволюции) представляют собой системы взаимосвязанных элементов, ориентированных на получение полезного для системы приспособительного результата.
В данной статье предложено обсуждение с привлечением кибернетических подходов таких проблем эволюции, как направленность эволюционных процессов и регулирующие механизмы видообразования.
2. Исторический экскурс
Одним из первых исследователей, указавших на роль в эволюции взаимодействий между органами, был В.С. Кирпичников (1940). В своей работе он продемонстрировал, как взаимодействия между органами, вызывающие адаптацию, закрепляются в процессе естественного отбора, а не в результате наследования приобретенных признаков по Ж. Ламарку. Уже первая попытка осознать роль взаимодействий в системах организма показала возможность устранения противоречий на основе системного подхода.
В 1948 году в результате исследований, проводимых в лаборатории П.К. Анохина, было обнаружено, что внешние факторы всегда оказывают влияние не на изолированный орган, а на функциональную систему в целом. На основании этого был сделан вывод о том, что эволюционируют и изменяются под воздействием внешних факторов всегда только функциональные системы, а морфологические трансформации внешних органов есть только отдаленное следствие этих изменений [4]. Одновременно с этим, Анохиным была продемонстрирована роль в естественном отборе гетерохронии √ неодновременной закладки и развития тканей и органов в эмбриогенезе.
Внутрисистемная гетерохрония √ это процесс сдвигания во времени закладки и изменения темпов развития отдельных фрагментов функциональной системы по сравнению с закладкой этих же фрагментов и для той же функциональной системы у предков. Внутрисистемная гетерохрония является одним из наиболее действенных средств организма, с помощью которых он отражает всякое длительное изменение в окружающей его обстановке; при условии, если эти изменения протекают на протяжении значительных геологических эпох [4]. Внутрисистемная гетерохрония является прямым следствием естественного отбора. Таким образом, оценивая время закладки отдельных фрагментов данной функциональной системы в сравнении с закладкой у предков в историческом плане, мы обращаемся к механизму возникновения данной функции в филогенезе.
В последствии на основании этих идей П.К. Анохиным был сформулирован принцип опережающего отражения действительности в функциональных системах. Согласно данному принципу в результате повторений каких-либо явлений в окружающей среде функциональные системы организмов, предназначенные для обслуживания этих явлений, приобретают свойство подготовиться по сигналу к еще только предстоящему событию [
4]. Для опережающего отражения действительности у животных существуют условные рефлексы, потребности и мотивации. С этой точки зрения нервную систему можно рассматривать, как ╚аппарат максимального и быстрейшего отражения последовательных и повторных явлений внешнего мира╩. У растений одним из вариантов опережающего отражения внешних условий можно считать реакцию развивающегося растения на действие внешней среды. Известно, что стрелолист (Sagittava sagittifilia) формирует различные листья под водой и в воздушной среде. Однако, фактором, определяющим развитие ╚подводных╩ листьев, служит не водная среда, а затемнение растения в момент развития. Таким образом, световые условия являются сигналом к развитию определенного типа листьев наиболее адекватно переносящих условия жизни, в которых будет существовать взрослое растение [5]. При опережающем отражении действительности пусковыми механизмами служит одно воздействие среды, а возникающие структуры, функции или реакции служат защитой от других воздействий среды. Не холод √ причина осеннего сбрасывания листьев, отлета птиц, линьки птиц и зверей, не засуха √ причина сбрасывания листьев растениями засушливого климата. Во многих случаях сигналом наступающих перемен, к которым надлежит приспособиться, служит изменение долготы дня. Возможно, в соответствии с принципом опережающего отражения формировалась способность первичных организмов активно реагировать последовательностью опережающих реакций на изменения во внешнем мире [4].Первым биологом-эволюционистом, подошедшим к проблемам эволюции с широким привлечением знаний и подходов кибернетики, был И.И. Шмальгаузен (1968). Он первым в мире развил взгляд на эволюцию как на регулируемый процесс. В 1939 и 1946 годах вышли его книги, посвященные возникновению регуляторных механизмов в процессе эволюции. И.И. Шмальгаузен показал, что отбор, способствуя образованию регуляторных механизмов индивидуального развития, меняет характер развития и создает устойчивые формы, остающиеся неизменными при неизменных условиях среды и способные меняться, как только условия среды изменились. В его работах показано, что гетерохрония, снижая зависимость развития систем друг от друга, повышает надежность организма в целом. И.И. Шмальгаузен впервые
предпринял попытку проанализировать взаимодействие организмов с биогеоценозом на основе обратной связи. Примерно в одно время с П.К.Анохиным И.И.Шмальгаузен пришел к мнению, что результат развития организмов и характер регуляции в ответ на воздействие внешней среды носит регуляторный характер. Внешние стимулы в виде сигналов воздействуют на организм, заставляя его подстраиваться к будущим изменениям среды. По сути, это было созвучно теории опережающего отражения действительности. Символично, что последняя и, к сожалению, незаконченная книга И.И. Шмальгаузена была посвящена месту кибернетики в теории эволюции.Анализируя проникновение идей кибернетики в эволюционное учение, невозможно не вспомнить одного из крупнейших ученых-кибернетиков А.А.Ляпунова. Рассматривая биологию с позиций кибернетики, он подчеркивал, что наиболее сложная проблематика теории эволюции √ это эволюция управляющих систем живой природы. А.А.Ляпунов высказал гипотезу о том, что появление новых структурных элементов на каком-либо ярусе организма приводит к формированию новых ярусов управляющих систем организма. При этом формирование высшего яруса управления легче всего может возникнуть в периоды неустойчивого режима [7].
Продолжением разработки эволюционных концепций с позиции кибернетики явилась работа ученого-физика и кибернетика В.Ф. Турчина (1977). В.Ф. Турчин сделал попытку рассмотреть биологическую эволюцию человека с позиций кибернетики. Для объяснения процессов эволюции В.Ф.Турчин предложил теорию ╚метасистемных переходов╩. Ее суть заключается в следующем: переход от нижних уровней системной иерархии к верхним происходит путем метасистемных переходов. При таких переходах формируются дополнительные механизмы управления группой систем более нижнего уровня. Примером метасистемных переходов, предложенным В.Ф. Турчиным, является следующая последовательность: управление положением = движение; управление движением = раздражимость (простой рефлекс); управление раздражимостью = (сложный) рефлекс; управление рефлексами = ассоциации (условный
рефлекс); управление ассоциациями = человеческое мышление; управление человеческим мышлением = культура. Теорию метасистемных переходов, по-видимому, можно рассматривать, как развитие гипотезы А.А. Ляпунова о процессе формирования новых ярусов в управлении организмом.Еще одним приложением кибернетики в анализе эволюционных процессов является их моделирование. Главными факторами, определяющими специфику моделирования эволюции, являются: ярко выраженная вариабельность биологических объектов, практически исключающая точное прогнозирование эволюции популяционных систем, и, как правило, невозможность экспериментальной проверки теоретической модели. В настоящее время моделирование в, смежной с эволюционной биологией, популяционной экологии рассматривается не столько как научный метод, сколько как искусство, где качество создаваемых моделей оценивается не только по соответствию научным критериям, но из соображений культуры и даже моды [9]. Очевидно, что данное мнение можно распространить и на моделирование эволюции. К сожалению, новаторские подходы в сфере моделирования эволюционных процессов возникают гораздо реже, чем, например, в физике, и зачастую не идут дальше заимствования аналитических средств из других областей знаний. Тем не менее, в результате развития идей моделирования в конце 80-х годов
XX века появились такие направления кибернетических исследований, как искусственная жизнь и эволюционная кибернетика. Примечательно, что изучением процессов эволюции стали заниматься физики и математики. Это ярко отражает многоплановость эволюционной теории. К сожалению, представители точных наук порой забывают о биологической составляющей этого феномена.Таким образом, если биология занимается в первую очередь изучением эволюции форм живых объектов, то кибернетика в первую очередь интересуется эволюцией строения и функционирования сложных систем, в том числе, таких как организм и популяция. Можно с уверенностью сказать, что кибернетика незаслуженно находится на периферии эволюционной биологии. Ранее уже неоднократно была продемонстрирована эффективность применения кибернетических подходов при изучении процессов эволюции.
3. Направленность эволюции
Общепринятая синтетическая теория эволюции плохо совмещается с концепцией о направленности эволюции. Однако еще в 1920 г. Н.И. Вавилов в своем законе гомологичных рядов наследственной изменчивости показал существование определенной направленности в изменчивости организмов и высказал мысль о возможности предсказания направления изменчивости у еще не изученных форм на основе анализа рядов изменчивости у родственных организмов. В последствии Л.С. Бергом была создана концепция номогенеза (эволюция на основе закономерностей) [10]. В предложенной концепции высказывалась мысль, что ╚изменчивость вовсе не безгранична; пределы ее могут быть на перед предсказаны╩. В настоящее время мнения о номогенезе полярны. С одной стороны концепция Л.С. Берга подвергается критике [11], с другой стороны продолжаются публикации отстаивающие позиции номогенеза [12]. По мнению Н.Н. Воронцова, главное позитивное в концепции направленной эволюции √ это представление о том, что в эволюции существуют запретные ходы, определенная направленность путей преобразования некоторых признаков и структур, что предает эволюции в какой-то степени направленный, канализируемый характер [13].
Говоря о номогенезе, критики и сторонники концепции имеют в виду, как правило, эволюцию признаков и структур. Однако, как было показано П.К. Анохиным, эволюционируют не отдельные органы, а функциональные системы содержащие их. Рассмотрим возможность направленности в эволюции с позиции теории функциональных систем.
Одно из важнейших свойств функциональных систем √ это их изоморфизм. Функциональные системы любого уровня организации имеют однотипную структуру и включают универсальные для разных систем компоненты: 1) полезный приспособительный результат; 2) рецепторы результата; 3) каналы обратной связи, передающие информацию от рецепторов в центральные структуры функциональной системы; 4) центральные структуры, являющиеся избирательным объединением нервных элементов различных уровней; 5) исполнительные соматические, вегетативные и эндокринные компоненты, включающие организованное и целенаправленное поведение [14]. Функциональные системы разных уровней и разных организмов могут иметь некоторые особенности, тем не менее, они всегда сохраняют изоморфизм их саморегулирующихся структур, обеспечивающих полезные приспособительные результаты, как для функциональной системы, так и для организма в целом [14].
Элементы функциональных систем неодинаково пластичны и, соответственно, неодинаково подвержены эволюционным изменениям. Наиболее консервативная часть функциональных систем √ результат действия и рецептор результата. Консервативность рецепторов обусловливает консервативный характер их сигнальной информационной активности. Соответственно консервативны и каналы, передающие эту информацию [14].
Пластичными свойствами в функциональных системах обладают управляющие структуры и периферические механизмы, обеспечивающие результат деятельности функциональной системы. Эти механизмы способны взаимозаменять и компенсировать деятельность друг друга. Показано, что после удаления одного легкого нагрузка в функциональной системе, обеспечивающей дыхательные функции, ложится не столько на второе легкое, сколько на сердце, почки и дыхательные свойства крови [4]. Поэтому при недостатке парциального давления кислорода в воздухе следует ожидать активацию дополнительных исполнительных механизмов для сохранения результата деятельности функциональной системы. Это основная причина выгодности мультифункциональных органов, способных в зависимости от необходимого результата выполнять ту или иную деятельность. Также для функциональных систем выгоден широкий охват исполнительных механизмов для обеспечения надежности получения результата. Например, такой специализированный орган, как крылья летучих мышей, выполняет функцию не только полета, но и схватывания добычи по принципу сачка, терморегуляции, продуцента витамина
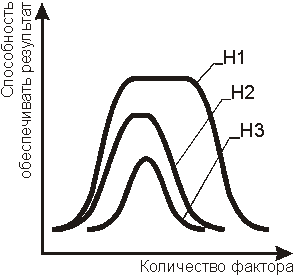
D, органа осязания [5].Свойства функциональных систем обуславливают их возможность сохранять постоянство результата при существенных изменениях условий их функционирования. Такие сохранительные способности тесно связаны с адаптационными и компенсаторными способностями организмов. На рисунке 1 показано, как в общем случае
способность функциональных систем организмов сохранять результат деятельности зависит от внешних условий [15]. График Н1 отражает способность системы сохранять результат своей деятельности за счет работы основных исполнительных механизмов системы. При работе основных исполнительных механизмов, служащих для адаптации к определенному фактору среды, их способность обеспечивать необходимый для организма результат находится в широком диапазоне изменений количества данного фактора. Графики Н2 и Н3 демонстрируют, как могут способствовать сохранению результата дополнительные, не основные исполнительные механизмы функциональной системы. Данная картина соответствует, приведенному ранее, примеру с удалением легкого, в котором основным исполнительным механизмом являются легкие, а дополнительными механизмами √ почки и дыхательные свойства крови. Очевидно, что разные исполнительные механизмы в силу своих особенностей не одинаково эффективно способны обеспечить необходимый результат. Другими словами, дополнительные исполнительные механизмы имеют разную готовность к адаптации в изменившихся условиях жизнедеятельности.
Рисунок
1. Способность обеспечивать постоянство результата исполнительными механизмами функциональной системы при изменениях условий среды (пояснения к рисунку даны в тексте).Это, по-видимому, обусловливает такое явление в эволюции, как относительная независимость темпов преобразования разных структур организма (гетеробатмия). При гетеробатмии быстро эволюционирующие органы компенсируют отставание медленно эволюционирующих в нужном направлении органов [16].
Известно, что чем активнее функционирует орган, тем меньше его изменчивость [5]. Такой орган находится под контролем стабилизирующей формы естественного отбора. Поэтому функционирование и строение структуры, обеспечивающей результат по графику Н1 (рис. 1), будет наименее изменчиво, а степень возможной компенсации результата структурами, соответствующими графикам Н2 и Н3, будет более изменчива. Учитывая наилучшие возможности компенсации при обеспечении результата у структуры Н2 и высокую изменчивость ее функциональности, можно сделать предположение, что данная структура при определенном направлении отбора будет эволюционировать наиболее быстро по сравнению со структурами Н1 и Н3. В итоге это приведет к усилению дополнительной функции у структуры Н2, и при продолжении давления направленного отбора произойдет смена главной функции. В свое время А.Н. Северцов, а позднее А.Л. Тахтаджян (1966) отмечали, что при переходе в новую адаптивную зону в первую очередь изменяются те системы организации, от которых непосредственно зависит адаптация к новым условиям среды. В дальнейшем если усиление дополнительной функции у структуры Н2 окажется недостаточным, чтобы обеспечить жизнедеятельность организма, то можно ожидать усиления дополнительной функции у Н3 и последовательного усиления функций у структур хоть как-то готовых к адаптации в изменившихся условиях. Это демонстрирует, что у организмов среди множества механизмов адаптации существуют более подготовленные для новых условий жизнедеятельности и, соответственно, в результате естественного отбора они разовьются раньше других.
Наименьшая скорость эволюции должна наблюдаться у рецептора результата. Поскольку он консервативен, то на него действует стабилизирующий отбор. Одним из древнейших механизмов, обеспечивающих консерватизм клеточных рецепторов, являются особые белки теплового шока (
HCP). Формирование этих белков присуще как растительным, так и животным клеткам различных таксонов [18]. Данные белки оказывают стабилизирующие неспецифическое действие на жизненно важные структуры клеток, в том числе на рецепторы [19, 20]. Они сохраняют до некоторой степени конформацию, т.е. трехмерную структуру, клеточных рецепторов и их функциональность даже при наличии в их структуре нарушений вызванных мутациями или повреждающими агентами. Также в функции этих белков входит: защита клеточных белков от теплового шока, кислородного голодания, химических повреждений и даже от атаки некоторых патогенов [20]. Если организм подвергнется, например, перегреву, часть белков может покинуть рецепторы для устранения перегрева. Оставленные ими рецепторы могут резко измениться, проявив все накопившиеся в них мутации. Если перегрев будет продолжаться в течение жизни нескольких поколений, то среди организмов резко увеличится изменчивость клеточных рецепторов, благодаря освобождению накопленного багажа мутаций. Существование такого явления подтвердили С.Резерфорд и С.Лундквист в своих экспериментах на дрозофилах (1998). Примечательно, что при данном явлении не возникают новые мутации, а проявляются уже существовавшие до этого мутации, другими словами обнаруживается скрытая изменчивость. Если проявление скрытой изменчивости будет способствовать выживанию организмов, то оно будет подхвачено естественным отбором. Таким образом, скрытая изменчивость также обусловливает существование структур, которые при определенном направлении отбора являются более готовыми для обеспечения жизненно важного результата.Возвращаясь к закону гомологических рядов наследственной изменчивости, действительно следует ожидать предсказуемости изменчивости у еще не изученных форм, но на основе выявления наиболее подготовленных путей изменчивости у родственных форм. Вышесказанное в значительной степени снимает противоречия между явлением направленности эволюции и синтетической теорией эволюции: эволюция идет на основе естественного отбора (селектогенез), но по наиболее подготовленному пути изменчивости. Наличие подготовленных путей изменчивости объясняет существование филогенетических параллелизмов √ одновременного возникновения в процессе эволюции сходств в структурах у разных организмов, живущих в одинаковых условиях. Наиболее часто параллелизмы наблюдаются у родственных организмов.
В настоящее время, пожалуй, еще не достаточно знаний, чтобы надежно прогнозировать усиление и смену функции у структур организмов в результате естественного отбора. Анализ гомологичных рядов √ единственный сейчас инструмент для прогноза.
Таким образом, в результате представленного анализа процессов эволюционного приспособления организма к окружающей среде с учетом теории функциональных систем показано, что у организмов существуют системы, более подготовленные к адаптации в новых условиях жизнедеятельности. Это позволяет допустить, что, зная эволюционные запреты, наиболее подготовленные пути изменчивости и направление отбора, можно с определенной вероятностью предсказать эволюцию.
4. Пути и регулирующие механизмы видообразования
Классическая дарвинская концепция представляет видообразование как постепенный, градуалистический процесс. Однако, Ч. Дарвин считал эволюцию процессом отнюдь не непрерывным, хотя и постепенным при переходе от одного вида к другому. Он сосредоточил основное внимание на исследовании дивергентной формы видообразования (кладогенез). Вместе с этим в своей работе Дарвин отразил и два других пути видообразования, получивших в последствии названия анагенез и стасигенез. Анагенез, по классификации Дж. Хаксли и Б. Ренш, является процессом прогрессивного развития группы, не обязательно связанный с ее распадением на боковые ветви [21]. Для значительного числа ископаемых форм с несомненностью доказано существование анагенетического пути развития. Эволюция человека была в основном анагенетическим процессом. Стасигенез √ это процесс длительного сохранения вида без прогрессивно направленного в каком-то направлении исторического развития и без дивергенции. В настоящее время выделяют еще одну форму происхождения таксонов √ симгенез, процесс слияния разных, ранее репродуктивно изолированных таксонов [13].
Наряду с представлением об эволюции как о постепенном процессе существует пунктуалистическая (скачкообразная) концепция видообразования. Сторонники этой концепции активно противопоставляют ее градуалистическому принципу эволюции. Одной из наиболее популяризованных теорий, возникшей на базе пунктуализма, явилась концепция ╚прерывистого равновесия╩ [22]. Согласно которой образование видов шло скачкообразно. Между образованием видов были длительные промежутки, в течение которых они не эволюционировали.
К настоящему моменту накоплено много фактического материала, подтверждающего существование, как градуализма, так и пунктуализма. При этом считается, что ╚обычный╩ путь видообразования, начинающийся с пространственной изоляции и завершающийся репродуктивной изоляцией, является градуалистическим. К пунктуализму относят ╚генетический╩ механизм видообразования, при котором репродуктивная изоляция возникает вследствие генетических и хромосомных изменений (например:
полиплоидия, горизонтальный перенос генов вирусами). Симгенез также является исключительно прерывистой формой видообразования [13].Проанализируем пути и механизмы видообразования с позиции кибернетических процессов управления и теории функциональных систем.
В соответствии с положениями, выдвинутыми И.И. Шмальгаузеном (1968), эволюция может рассматриваться как автоматически регулируемый или, другими словами, саморегулирующийся процесс (рис. 2). В качестве регулирующего механизма выступает экосистема. Объектом управляемых изменений является популяция, как наименьшая единица эволюции. Связь между экосистемой и популяцией осуществляется в двух направлениях. Прямая информация об экосистеме передается в популяцию через апробированных особей в виде их генотипа. Обратная информация в экосистему возвращается в виде фенотипа потомства выживших особей. Фенотип особи проходит в течение своего жизненного цикла испытания в экосистеме. В процессе испытаний часть особей гибнет, а другая часть доживает до оставления потомства. Естественный отбор является выражением результатов сложной системы контроля в экосистеме. Отбор служит основным механизмом преобразования обратной информации в экосистеме [6].

Рисунок
2. Автоматически регулируемая эволюция (И.И. Шмальгаузен, 1968)Изобразим описанную схему эволюции на основе положений кибернетической теории об управляющих системах [23, 24]. Управляющая система √ объект способный воспринимать, перерабатывать и выдавать информацию. Выдаваемая информация характеризует поведение управляющей системы. В качестве управляющей системы выступает, несомненно, экосистема (рис. 3). Объект управления экосистемы √ популяция. Параметром управления является фенотип. Экосистема выдает информацию в виде генотипа апробированных особей. По каналам обратной связи передается информация о состоянии объекта управления (популяции), о количественном и качественном составе популяции. Обратная связь о состоянии параметра управления (фенотипе) содержит информацию о его состоянии (морфофизиологических особенностях). Сравнение представлений И.И. Шмальгаузена и схемы, созданной нами с использованием теории управляющих систем, обнаруживает их значительное сходство. Вместе с этим показано, что в экосистему помимо информации о морфофизиологических свойствах фенотипа передается также информация о качественном и количественном составе популяции.
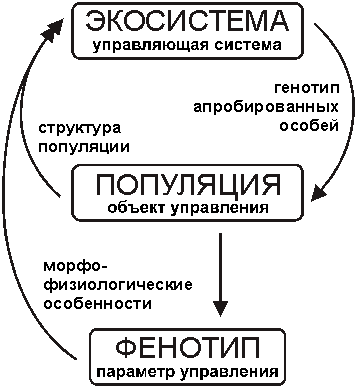
Рисунок
3. Управляющая система в эволюцииВ обеих схемах функция естественного отбора заключается в испытании организмов на выживаемость в экосистеме и в удалении организмов, не прошедших испытаний. Очевидно, что разнообразие фенотипов, создающее поле деятельности для естественного отбора, обусловлено генетической
изменчивостью. Проанализируем механизмы, формирующие изменчивость фенотипов в саморегулирующемся процессе эволюции в соответствии с представлением об управляющих системах.В саморегулирующихся управляющих системах действует ╚золотое правило╩ саморегуляции: само отклонение параметра управления от заданного значения запускает систему, чтобы скорректировать параметр управления [14]. Применительно к эволюции под параметром управления здесь подразумевается фенотип, а ╚заданное значение╩ обозначает условия в экосистеме.
Это правило продолжает известный принцип Ле Шателье о внутреннем, встроенном управлении в системах. Согласно этому принципу, если на любую систему, находящуюся в стационарном режиме, подействует внешнее возмущение, то в ней произойдут такие изменения, которые уменьшат результат его действия.Внутреннее управление обеспечивается в саморегулирующихся системах тремя основными способами управления: пропорциональным, дифференциальным и интегральным [25]. С помощью этих трех способов или законов управления управляющая система (экосистема) формирует управляющее воздействие на объект управления (популяцию). Проанализируем роль законов управления применительно к эволюционному процессу адаптации организмов к изменившимся условиям экосистемы.
Первый закон (пропорциональный) - управление по параметру отклонения. Согласно закону, управление происходит пропорционально отклонению получаемого результата от ожидаемого (рис. 4). Под получаемым результатом подразумеваются морфофизиологические особенности фенотипов, возникающих как результат размножения особей, апробированных естественным отбором. Ожидаемым результатом являются такие морфофизиологические особенности фенотипов, которые дадут возможность организмам адекватно адаптироваться в изменившихся условиях экосистемы. В связи с тем, что в соответствии с данным законом изменения происходят пропорционально и постепенно, то можно предположить, что данный закон отражает усиление главной функции у структур ответственных за обслуживание тех параметров экосистемы, которые изменились. Например, при устойчивом увеличении средней температуры в экосистеме с теплым климатом пропорционально усилится функция теплоотводящих структур организма. Поскольку при усилении главной функции необходимые структуры уже существуют, то, очевидно, что данный процесс идет за счет постепенного развития и совершенствования ответственных структур. Усиление главной функции происходит либо в результате постепенного изменения составляющих такую структуру единиц, либо при увеличении числа компонентов
внутри структуры. Пример первого рода √ усиление функции мышечного сокращения в результате замещения гладкой мускулатуры поперечно-полосатой. Пример второго рода √ развитие млечных желез у млекопитающих, идущее по пути значительного увеличения числа отдельных долек, формирующих вместе более мощную железу [5].
Рисунок
4. Первый закон управления (пропорциональный). График F √ изменение какого-либо фактора в экосистеме, график 1 √ усиление главной функции у структуры, ответственной за адаптацию к данному фактору.Согласно второму закону управления, в системах существует регуляция пропорциональная скорости отклонения получаемого результата от ожидаемого (дифференциальный закон) (рис. 5). Данный закон отражает включение у организмов готовых компенсаторных и дополнительных исполнительных механизмов в функциональных системах при изменении условий в экосистеме. При этом изменчивость структуры данных механизмов, как не основных, в популяции относительно высока. Среди механизмов существуют более готовые к обеспечению адаптации к новым условиям (наиболее подготовленные пути изменчивости), которые и подхватываются естественным отбором. Однако количество подготовленных путей ограниченно, поэтому их вклад в изменчивость фенотипа после увеличения будет уменьшаться. График, отражающий суммарную изменчивость фенотипа в результате управления по пропорциональному и дифференциальному законам, демонстрирует, что на переходном этапе экосистемы будут существовать фенотипы
с опережающей готовностью к новым условиям.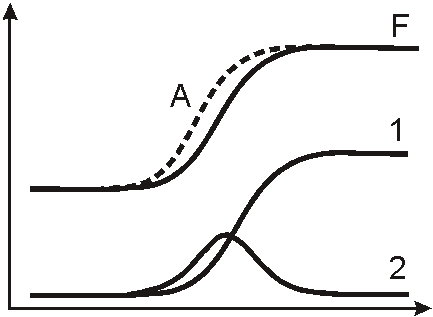
Рисунок
5. Второй закон управления (дифференциальный). График 2 √ включение наиболее подготовленных путей изменчивости для адаптации к изменению фактора F; график A √ суммарная изменчивость в результате работы пропорционального и дифференциального законов; графики F и 1 как на рис. 4.Процесс естественного отбора при управлении по пропорциональному и дифференциальному законам совпадает с процессом, описанным И.И. Шмальгаузеном (1968, с. 101). ╚В первой фазе естественного отбора положительных вариантов происходит их накопление при очень небольшом коэффициенте селекции. По мере нарастания концентрации скорость естественного отбора сначала быстро возрастает
до максимума, лежащего на уровне 0,4-0,5 концентрации отбираемого фена, а затем несколько медленно падает┘ В заключительной фазе завоевания всей популяции, т.е. становления новой формы, естественный отбор замедляется и переходит в стабилизирующую форму с преобладающей элиминацией неблагоприятных уклонений от этой нормы.╩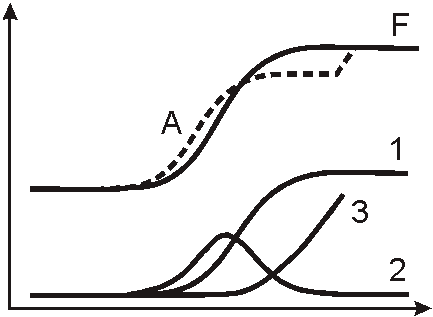
Рисунок
6. Третий закон управления (интегральный). График 3 √ процесс накопления мутаций в результате стресса; графики A, F, 1, 2 как на рисунках 4 и 5.Третий закон управления реализуется, в случае если механизмы, действующие по первым двум законам, не справляются, т.е. не обеспечивают адекватную адаптацию организма к окружающим условиям. Согласно третьему закону, воздействие осуществляется пропорционально общей сумме отклонения получаемого результата от ожидаемого во времени (интегральный закон) (рис. 6). Применительно к эволюции, очевидно, что этому закону управления соответствует накопление мутаций. Интегральный закон проявляется в управлении, если морфофизиологические свойства фенотипа остаются неудовлетворительными в результате действия пропорционального и дифференциального законов управления. Мутации увеличивают изменчивость структур и органов у организмов и расширяют поле возможностей для естественного отбора. В итоге может появиться фенотип, имеющий адаптивные приспособления к новым условиям.
Существование интегрального закона управления требует наличия механизма, запускающего мутации для увеличения изменчивости структур при недостаточной приспособленности организма к внешним условиям. Мы полагаем, что таким механизмом является стресс - неспецифическая адаптационная реакция, возникающая при сильных воздействиях на организм.
При встрече организма с тем или иным экстремальным агентом, сила и длительность воздействия которого выводят организм за пределы условий обычной жизнедеятельности, усиливаются катаболические процессы, возрастает количество окисленных или частично окисленных активных форм кислорода, в том числе свободных радикалов, происходит активация перекисного окисления липидов. Активация перекисного окисления наблюдается при стрессах самой разнообразной природы, вызванных воздействием ультрафиолетовой радиации, высокой и низкой температур, гипероксии и гипоксии, физических нагрузок и их отсутствием, интоксикаций, эмоциональных и психических напряжений и т.п. [26]. Продукты перекисного окисления являются высоко реакционными и, попадая в клеточные ядра, вызывают точковые мутации и хромосомные абберации. В организмах существуют антиоксидантные, стресслимитирующие системы, которые ограничивают эту активацию, препятствуя ее продолжению, и удерживают образование продуктов перекисного окисления в пределах, близких к оптимуму жизнедеятельности [27]. Продукты перекисного окисления всегда присутствуют в организме. Их мутагенное воздействие нейтрализуется репарационными системами клеток. Функция таких систем заключается в исправлении химических повреждений в молекулах ДНК, т. е. в препятствовании появлению мутаций. Однако при тяжелых и продолжительных воздействиях, когда исчерпываются мощности защитных систем, количество продуктов перекисного окисления значительно возрастает, что приводит к необратимым повреждениям и разрушениям биомембран, клеток, тканей. При полном исчерпании ресурсов стресслимитирующих защитных систем (например, у лососевых после нереста) резко возрастает количество токсических продуктов перекисного окисления, ведущее к необратимому нарушению жизненных функций и к гибели организма [28]. Таким образом, стресс и его продукты оказывают повреждающее и мутагенное действие на организмы при сильных воздействиях и недостаточной приспособленности организма к изменившимся внешним условиям.
Выраженность стресса, характеризующая степень повреждения организма, зависит от величины внешних воздействий на организм. При незначительной величине внешних воздействий, когда организм находится в благоприятных условиях, стресс не происходит, а если и происходит, то стресслимитирующие и репарационные механизмы организма полностью устраняют
его последствия. При чрезмерном уровне внешних воздействий повреждения и мутации, возникающие в результате стресса, будут иметь патологический характер и вызовут быструю гибель организма. Между незначительными воздействиями и чрезмерными находится коридор таких величин воздействия, при которых стресс ведет к накоплению мутаций в организме, но при этом повреждения организма незначительны, и он способен дожить до момента передачи своего генотипа потомству. Таким образом, существует определенный коридор величин внешних воздействий на организм, и только при нахождении в нем у организмов возникают мутации и передаются потомству.Говоря о стрессе, часто подразумевают его существование только у животных с высоким уровнем организации, обладающих центральной нервной системой и механизмами регуляции гомеостаза. Это связано с исследованиями Г. Селье, который считал типичными такие проявления стресса, как активация гипофизарно-адреналовой системы, инволюция и деструкция тимико-лимфотической системы, возникновение язв желудка и кровотечений. Однако, как показали более поздние исследования, неспецифические реакции на внешние воздействия наблюдаются, в том числе, на уровне клеток и могут рассматриваться как клеточный стресс [19, 29]. Стресс в главных чертах присущ всем
живым организмам. Разумеется, что его механизмы совершенствовались по мере усложнения и повышения организации живых существ.Итак, изменение условий в экосистеме включает механизмы видообразования. Если в экосистеме условия стабильны, то механизмы видообразования не работают, и естественный отбор направлен на сохранение уже достигнутых приспособлений. Такую форму отбора называют стабилизирующей [30]. При стабилизирующем отборе новые виды организмов не образуются.
В 1970-х годах были открыты уникальные экосистемы √ ╚черные курильщики╩, гидротермальные сероводородные источники, расположенные на больших глубинах в океанах. Свойства экосистемы, окружающей ╚черных курильщиков╩, не менялись, по крайней мере, не менее 400 млн. лет. Палеонтологические данные показали, что в прошлом состав продуцентов этих экосистем (бактериальных сообществ) практически не изменился, а гетеротрофное население сформировалось за счет эмигрантов из ╚нормальных╩ биотопов. Вестиментиферы, по-видимому, единственный специализированный многоклеточный организм этих гидротермальных экосистем. У вестиментифер не установлено за последние 400 млн. лет признаков эволюционного развития. В гидротермальных оазисах найдены также представители других групп животных, считавшихся вымершими более сотни миллионов лет назад [31]. ╚Черные курильщики╩ демонстрируют отсутствие прогрессивной эволюции в стабильной среде.
Реликтовые формы организмов, сохранившие неизменными основные особенности строения на протяжении огромных промежутков времени, существуют не только на ╚черных курильщиках╩. Так, морское плеченогое лингула (
Lingula) существует как вид более 500 млн. лет, моллюск Neopilina не менее 400 млн. лет, мечехвост (Limulus) около 400 млн. лет, кистеперая рыба латимерия (Latimeria) около 300 млн. лет. Число примеров можно увеличить. В большинстве случаев сохранение видов неизменными стало возможно благодаря стабильности основных компонентов среды [5]. Особенно много реликтовых организмов в некоторых участках Мирового океана и в тропиках, где условия существования остаются стабильными на протяжении десятков миллионов лет.По-видимому, в стабильной среде, когда не включены механизмы видообразования, эволюция носит финалистический характер. Другой, возможной причиной появления стабильной, не эволюционирующей формы в некоторых случаях может явиться нарушение у неё механизмов видообразования. Таким нарушением, возможно, является высокая устойчивость к стрессам или чрезмерная мощность репарационных механизмов. Данное предположение допускает экспериментальную проверку
.Представленный анализ, безусловно, является несколько идеализированным. Например, за рамками рассмотрения остались такие факторы, как конкуренция видов за ресурсы, их взаимодействия, экспоненциальный рост численности популяций и т.д. Тем не менее, предложенные подходы допускают учет этих факторов. Описанная схема эволюционных процессов может быть использована для компьютерного моделирования эволюции.
Таким образом, выполненный анализ эволюции, как саморегулирующегося процесса, сделал возможным выделение механизмов, обеспечивающих ее внутреннее управление. В качестве механизмов, определяющих изменчивость организмов, предложены: усиление главной функции органов, активация наиболее подготовленных путей изменчивости, стресс √ как поставщик мутационной изменчивости. Показано, что данные механизмы начинают работать только при достаточных изменениях окружающей среды. Вместе с этим чрезмерные изменения среды приведут к вымиранию организмов. В стабильной окружающей среде эволюция носит финалистический характер.
Заключение
В результате анализа процессов эволюционного приспособления организма к окружающей среде с учетом теории функциональных систем показано, что у организмов существуют системы неодинаково подготовленные к адаптации в новых условиях. Это обусловливает существование наиболее подготовленных путей изменчивости. Знание эволюционных запретов, наиболее подготовленных путей изменчивости и направление отбора допускает предсказание с определенной вероятностью течения эволюции.
Подход к эволюции, как к саморегулирующейся системе, продемонстрировал, что эволюция, как процесс, идет непрерывно, но неравномерно, ей свойственны как скачки, так и периоды длительного сохранения вида без изменений. Ход эволюции организмов определяется свойствами окружающей их экосистемы
. В качестве механизмов, обусловливающих изменчивость организмов, предложены: усиление главной функции органов, активация наиболее подготовленных путей изменчивости, стресс √ как поставщик мутационной изменчивости. Представленная схема эволюционных процессов может быть использована для математического моделирования эволюции.Результаты нашего анализа не противоречат положениям современной теории эволюции. Вместе с этим они позволили увидеть механизмы, лежащие в основе эволюционных изменений и, в некоторой степени, обобщить их. Предложенный материал продемонстрировал, что кибернетическая проблематика эволюции чрезвычайно широка и разнообразна. Можно с уверенностью сказать, что кибернетика является необходимой составной частью эволюционной биологии и в будущем
может служить основой для нового эволюционного синтеза.Список литературы
| цкюбмюъ | днйкюдш | наясфдемхе | |